
William G. Kaelin Jr., Sir Peter J. Ratcliffe y Gregg L. Semen descubrieron cómo las células pueden detectar y adaptarse a la disponibilidad cambiante de oxígeno. Identificaron la maquinaria molecular que regula la actividad de los genes en respuesta a los niveles variables de oxígeno.
Los descubrimientos fundamentales de los premios Nobel de este año revelaron el mecanismo para uno de los procesos adaptativos más esenciales de la vida. Establecieron las bases para nuestra comprensión de cómo los niveles de oxígeno afectan el metabolismo celular y la función fisiológica. Sus descubrimientos también han allanado el camino para nuevas estrategias prometedoras para combatir la anemia, el cáncer y muchas otras enfermedades.
Oxígeno en el centro del escenario
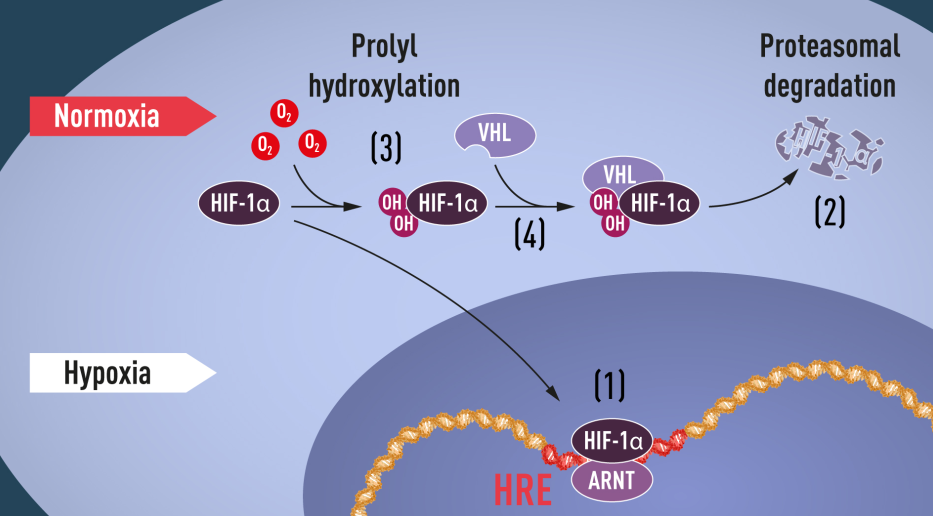 |
| Al presentarse niveles de oxígeno bajos (hipoxia), HIF-1a protege la degradación y se acumula en el núcleo, donde se asocia con ARNT y se une a secuencias de ADN específicas (HRE) en genes regulados por hipoxia (1) A niveles normales de oxígeno, el proteasoma degrada rápidamente el HIF-1a (2). El oxígeno regula el proceso de degradación mediante la adición de grupos hidroxilo (OH) a HIF-1a (3). La proteína VHL puede reconocer y forman un complejo con HIF-1a que conduce a su degradación de una manera dependiente del oxígeno (4). |
Otto Warburg, el ganador del Premio Nobel de Fisiología o Medicina de 1931, reveló que esta conversión es un proceso enzimático. Durante la evolución, se volvió un mecanismo para asegurar un suministro suficiente para los tejidos y células. El cuerpo carotídeo, adyacente a los vasos sanguíneos grandes en ambos lados del cuello, contiene células especializadas que detectan los niveles de oxígeno en la sangre. El Premio Nobel de Fisiología o Medicina de 1938 Corneille Heymans descubrió como la detección del oxígeno en la sangre a través del cuerpo carotídeo controla nuestra respiración por medio de la comunicación directamente con el cerebro.
HIF entra en escena
Además de la adaptación rápida controlada por el cuerpo carotídeo para reducir los niveles de oxígeno (hipoxia), existen otras adaptaciones fisiológicas fundamentales. Una respuesta fisiológica clave a la hipoxia es el aumento en los niveles de la hormona eritropoyetina (EPO), que conduce a una mayor producción de células sanguíneas rojas. La importancia del control hormonal de la eritropoyesis ya se conocía a principios del siglo XX, pero la forma en que este proceso fue controlado por O2 siguió siendo un misterio.
Gregg Semenza estudió el gen EPO y cómo está regulado por los niveles variables de oxígeno. Al utilizar ratones modificados genéticamente, se demostró que segmentos específicos de ADN ubicados junto al gen EPO median la respuesta a la hipoxia. Sir Peter Ratcliffe también estudió la regulación dependiente de O2 del gen EPO, y ambos grupos de investigación encontraron que el mecanismo de detección de oxígeno estaba presente en prácticamente todos los tejidos, no solo en las células renales donde normalmente se produce EPO. Estos fueron hallazgos importantes que demostraron que el mecanismo era general y funcional en muchos tipos de células diferentes.
Semenza deseaba identificar los componentes celulares que median esta respuesta. En las células hepáticas cultivadas descubrió un complejo de proteínas que se une al segmento de ADN identificado de una manera dependiente del oxígeno. Llamó a este: factor inducible por la hipoxia completa (HIF). Comenzaron los esfuerzos extensivos para purificar el complejo HIF, y en 1995, Semenza pudo publicar algunos de sus hallazgos clave, incluida la identificación de los genes que codifican HIF. Se descubrió que el HIF constaba de dos proteínas de unión al ADN diferentes, denominadas factores de transcripción, ahora llamados HIF-1a y ARNT. Ahora los investigadores podrían comenzar a resolver el rompecabezas, permitiéndoles comprender qué componentes adicionales estaban involucrados y cómo funciona la maquinaria.
VHL: un compañero inesperado
Cuando los niveles de oxígeno son altos, las células contienen muy poco HIF-1a. Sin embargo, cuando los niveles de oxígeno son bajos, la cantidad de HIF-1 aumenta para que pueda unirse y así regular el gen EPO y otros genes con segmentos de ADN que se unen a HIF (Figura 1). Varios grupos de investigación mostraron que HIF-1a, que normalmente se degrada rápidamente, está protegido de la degradación en la hipoxia. A niveles normales de oxígeno, la máquina acelular llamada proteasoma, reconocida por el Premio Nobel de Química 2004 a Aaron Ciechanover, Avram Hershko e Irwin Rose, degrada el HIF-1a. En tales condiciones, se agrega un péptido pequeño, ubiquitina, a la proteína HIF-1. La ubiquitina funciona como una etiqueta para proteínas destinadas a la degradación en el proteasoma. Cómo la ubiquitina se une al HIF-1 en una forma dependiente de oxígeno sigue siendo una pregunta central.La respuesta vino de una dirección inesperada. Casi al mismo tiempo que Semenza y Ratcliffe estaban explorando la regulación del gen EPO, el investigador del cáncer William Kaelin, Jr. estaba investigando un síndrome hereditario, la enfermedad de von Hippel-Lindau (enfermedad de VHL). Esta enfermedad genética conduce a un riesgo dramático de ciertos cánceres en familias con mutaciones de VHL heredadas. Kaelin demostró que el gen VHL codifica una proteína que previene la aparición del cáncer. Kaelin también mostró que las células cancerosas que carecen de un gen VHL funcional expresan niveles anormalmente altos de genes regulados por hipoxia; pero que cuando el gen VHL se reintrodujo en las células cancerosas, se restablecieron los niveles normales. Esta fue una pista importante que muestra que la BVS estuvo de alguna manera involucrada en el control de las respuestas a la hipoxia. Se obtuvieron pistas adicionales de varios grupos de investigación que muestran que VHL es parte de un complejo que etiqueta las proteínas con ubiquitina, marcándolas para la degradación en el proteasoma. Ratcliffe y su grupo de investigación hicieron un descubrimiento clave: demostrar que la BVS puede interactuar físicamente con el HIF-1a y es necesaria para su degradación a niveles normales de oxígeno. Esto vincula VHL a HIF-1a de manera concluyente.
El oxígeno reduce el equilibrio
Muchas piezas se habían acomodado, pero aún faltaba comprender cómo los niveles de O2 regulaban la interacción entre VHL y HIF-1a. La búsqueda se centró en una porción específica de la proteína HIF-1 conocida por ser importante para la degradación dependiente de VHL , y tanto Kaelin como Ratcliffe sospecharon que la clave para la detección de O2 residía en algún lugar de estas proteínas. En 2001, en dos artículos publicados simultáneamente, demostraron que bajo niveles normales de oxígeno, los grupos hidroxilo se colocan en dos posiciones específicas en HIF-1a. Esta modificación de la proteína, llamada prolil hidroxilación, permite que VHL reconozca y se una al HIF-1a y, por lo tanto, explica cómo los niveles normales de oxígeno controlan la rápida degradación del HIF-1 con la ayuda de enzimas sensibles al oxígeno (las llamadas prolil hidroxilasas). Investigaciones posteriores de Ratcliffe y otros identificaron las prolil hidroxilasas responsables. También se demostró que la función de activación de genes de HIF-1a estaba regulada por hidroxilación dependiente de oxígeno. Los galardonados con el Premio Nobel ya habían dilucidado el mecanismo de detección de oxígeno y habían demostrado cómo funciona.
Oxígeno forma fisiología y patología
Gracias al innovador trabajo de estos Nobel Laureados, sabemos mucho más acerca de cómo los diferentes niveles de oxígeno regulan los procesos fisiológicos fundamentales. La detección de oxígeno permite a las células adaptar su metabolismo a niveles bajos de oxígeno: por ejemplo, en nuestros músculos durante el ejercicio intenso. Otros ejemplos de procesos adaptativos controlados por la detección de oxígeno incluyen la generación de nuevos vasos sanguíneos y la producción de glóbulos rojos. Nuestro sistema inmunológico y muchas otras funciones fisiológicas también están ajustadas por la detección de O2. Incluso se ha demostrado que la detección de oxígeno es esencial durante el desarrollo fetal para controlar la formación normal de vasos sanguíneos y el desarrollo de la placenta.
La detección de oxígeno es fundamental para una gran cantidad de enfermedades. Por ejemplo, los pacientes con insuficiencia renal crónica a menudo sufren de anemia severa debido a la disminución de la expresión de EPO. EPO es producido por células en el riñón y es esencial para controlar la formación de glóbulos rojos, como se explicó anteriormente. Además, la máquina con oxígeno regulado tiene un papel importante en el cáncer. En tumores tumores, la maquinaria regulada por oxígeno se utiliza para estimular la formación de vasos sanguíneos y remodelar el metabolismo para la proliferación efectiva de células cancerosas. Intensos esfuerzos continuos en laboratorios académicos y compañías farmacéuticas ahora se centran en desarrollar medicamentos que puedan interferir con diferentes estados de enfermedad activando o bloqueando la maquinaria de detección de oxígeno.





0 Comments
Publicar un comentario